

 English
English Bahasa Melayu
Bahasa Melayu Bahasa Indonesia
Bahasa Indonesia Tiếng Việt
Tiếng Việt ไทย
ไทย
本翻譯僅作學術交流用,無商業意圖,請勿轉載,如有疑議問請來信
研究顯示,遠紅外線(IFR)治療對糖尿病小鼠的缺血誘導血管生成具有顯著效果。實驗中,IFR治療不僅提高了缺血肢體的血流恢復,也增強了內皮祖細胞(EPC)的功能。這項發現為糖尿病相關血管疾病的治療提供了新的可能性,展示了遠紅外線應用於心血管疾病治療的潛力。
遠紅外線治療促進糖尿病小鼠的缺血誘導血管生成,並恢復高血糖抑制的內皮祖細胞功能。
Far infra-red therapy promotes ischemia-induced angiogenesis in diabetic mice and restores high glucose-suppressed endothelial progenitor cell functions
Po-Hsun Huang, Jaw-Wen Chen, Chih-Pei Lin, Ying-Hwa Chen, Chao-Hung Wang, Hsin-Bang Leu & Shing-Jong Lin
Cardiovascular Diabetology volume 11, Article number: 99 (2012) Cite this article
https://cardiab.biomedcentral.com/articles/10.1186/1475-2840-11-99
摘要
背景
遠紅外線(IFR)治療已被證明對心血管系統產生有益效果,但IFR對內皮祖細胞(EPC)和EPC相關的血管生成的效果尚不清楚。我們假設IFR輻射可以通過增強EPC的功能和歸巢過程,恢復糖尿病小鼠缺血性後肢的血流恢復。
材料和方法
在糖尿病發病4週後,以链脲佐菌素(STZ)誘導的糖尿病小鼠中誘導單側後肢缺血,然後將其分為對照組和IFR治療組(每組6隻)。後者小鼠被放置在34°C的IFR乾燥桑拿浴中,每天30分鐘,持續5週。
結果
多普勒灌注成像顯示,熱療法組的缺血肢體/正常側血灌注比率明顯高於對照組,並且在IFR治療組中看到明顯更大的毛細血管密度。流式細胞分析顯示,無論是否接受IFR治療,糖尿病小鼠在缺血手術後EPCs(Sca-1+/Flk-1+)的動員受損(每組6隻)。然而,與對照組相比,接受IFR輻射的糖尿病小鼠的骨髓源EPCs分化為被定義為GFP+/CD31+雙陽性細胞的內皮細胞,在缺血組織周圍的血管中明顯增加。在體外研究中,用IFR輻射治療的培養EPCs顯著增強了高葡萄糖損傷的EPC功能,抑制了高葡萄糖誘導的EPC衰老並減少了H2O2產生。裸鼠在高葡萄糖培養基中接受IFR治療的人類EPCs顯示,與未接受IFR治療的鼠類相比,缺血肢體的血流恢復明顯改善。IFR治療促進了STZ誘導的糖尿病小鼠的血流恢復和新血管形成。
結論
IFR治療的施用促進了STZ誘導的糖尿病小鼠的側向流量恢復和新血管形成,這些有益效果可能來自於增強EPC功能和歸巢過程。
Abstract
Background
Far infra-red (IFR) therapy was shown to exert beneficial effects in cardiovascular system, but effects of IFR on endothelial progenitor cell (EPC) and EPC-related vasculogenesis remain unclear. We hypothesized that IFR radiation can restore blood flow recovery in ischemic hindlimb in diabetic mice by enhancement of EPCs functions and homing process.
Materials and methods
Starting at 4 weeks after the onset of diabetes, unilateral hindlimb ischemia was induced in streptozotocine (STZ)-induced diabetic mice, which were divided into control and IFR therapy groups (n = 6 per group). The latter mice were placed in an IFR dry sauna at 34°C for 30 min once per day for 5 weeks.
Results
Doppler perfusion imaging demonstrated that the ischemic limb/normal side blood perfusion ratio in the thermal therapy group was significantly increased beyond that in controls, and significantly greater capillary density was seen in the IFR therapy group. Flow cytometry analysis showed impaired EPCs (Sca-1+/Flk-1+) mobilization after ischemia surgery in diabetic mice with or without IFR therapy (n = 6 per group). However, as compared to those in the control group, bone marrow-derived EPCs differentiated into endothelial cells defined as GFP+/CD31+ double-positive cells were significantly increased in ischemic tissue around the vessels in diabetic mice that received IFR radiation. In in-vitro studies, cultured EPCs treated with IFR radiation markedly augmented high glucose-impaired EPC functions, inhibited high glucose-induced EPC senescence and reduced H2O2 production. Nude mice received human EPCs treated with IFR in high glucose medium showed a significant improvement in blood flow recovery in ischemic limb compared to those without IFR therapy. IFR therapy promoted blood flow recovery and new vessel formation in STZ-induced diabetic mice.
Conclusions
Administration of IFR therapy promoted collateral flow recovery and new vessel formation in STZ-induced diabetic mice, and these beneficial effects may derive from enhancement of EPC functions and homing process.
引言
血管生成,即從既存的血管結構中血管的萌發,是缺血的生理反應,但通常無法產生足夠的便道流量來挽救缺血器官[1]。在缺血的動物模型中,越來越多的證據表明,通過重組蛋白或基因轉移,施用血管生成生長因子可以通過新血管生成增加營養灌注並減少與缺血有關的器官損傷[2]。然而,有臨床證據表明,某些患者與健康的實驗動物不同,組織缺血後未能形成便道循環,並似乎對外源生長因子不敏感[3]。糖尿病患者與內皮功能障礙有關,並且在心臟和周邊組織的缺血反應中減少了便道血管的形成能力[4-6]。
有力的證據表明,成人的新血管生成不僅是當地內皮細胞(血管生成)的增殖結果,還涉及骨髓衍生的循環內皮祖細胞(EPCs)進行血管生成的過程[7]。然而,有證據表明,具有心血管風險因素的患者的循環EPCs的數量和功能都減少了[8]。我們最近的研究還表明,長期暴露於高葡萄糖可能通過一氧化氮(NO)相關機制增強細胞衰老,減少EPCs的細胞數量和功能能力[9]。
近期證據顯示,2型糖尿病(DM)患者的循環EPCs水平和動脈僵硬度與其血糖控制密切相關[10]。這些發現為糖尿病患者高血糖相關的血管併發症提供了潛在治療目標的合理依據。遠紅外線(IFR)輻射是一種不可見的電磁波,其特徵波長介於5.6和1000μm之間,可以通過皮膚中的熱感受器感知為熱[11,12]。最近的研究表明,IFR治療對心血管系統產生有益效果。 IFR輻射改善心臟病患者的心室心律失常和內皮功能,並增強血液透析患者的動靜脈瘻的通流量和通暢度[13-15]。此外,IFR治療促進各種動物模型的微血管血流和血管生成[16,17]。雖然臨床研究已經表明IFR輻射可以對心血管系統產生有益效果,但IFR治療對循環EPCs和糖尿病的多方面效果仍不清楚。因此,我們假設IFR輻射可以改善糖尿病小鼠組織缺血後的血流恢復,並通過增加NO生物可用性提高EPCs的功能能力。
材料和方法
動物
6-8周大的小鼠由台灣國家實驗動物中心購得(FVB小鼠)。通過每天腹腔注射柑橘酸緩衝液中的链脲佐菌素(STZ)(40 mg/kg)連續5天,以誘導FVB小鼠的1型糖尿病模型[18]。只有在第一次STZ注射後14天血糖濃度超過250 mg/dl並出現明顯糖尿時,才認為小鼠患有糖尿病。糖尿病的持續性在研究結束時確定。所有小鼠均放置在微隔離籠中,按12小時的白天/夜晚週期飼養。該調查符合美國國立衛生研究院(NIH出版物1996)發布的實驗動物護理和使用指南。國立陽明大學(台北,台灣)的機構動物護理委員會批准了所有涉及動物的實驗程序和方案,並符合ARRIVE指南[19]的規定。
小鼠缺血性後肢模型
八周大的雄性野生型小鼠和STZ誘導的糖尿病小鼠通過切除右側股動脈接受單側後肢缺血手術,如先前所述[20]。在手術後的2週內,STZ誘導的糖尿病小鼠每天兩次接受30分鐘的局部IFR治療,而野生型和糖尿病對照小鼠則每天兩次放在34°C的加熱板上30分鐘,以避免本研究中的熱效應。簡單地說,小鼠通過腹腔注射氯胺酮(100 mg/kg)和地塞米松(10 mg/kg)進行麻醉。通過確保疼痛的捏刺激(鈍鉗)在後爪、前爪和耳朵上不引起任何運動反射來檢查麻醉的深度。股動脈的近端和遠端部分被結紮。使用激光多普勒灌注成像系統(Moor Instruments Limited,英國德文)測量手術前後及每週隨訪的後肢血灌注。為了避免環境光和溫度的影響,結果表示為右側(缺血)與左側(非缺血)肢體的灌注比例。
測量缺血肢體的毛細血管密度和氧化應激
手術後4週,通過靜脈注射氯胺酮對小鼠進行安樂死。小心地移除股骨,並將缺血的大腿肌肉嵌入石蠟中。切片(5μm)去蠟化並孵育與抗鼠CD31的大鼠單克隆抗體(克隆MEC13.1,BD PharMingen,聖地亞哥,CA)。使用生物素-辣根過氧化酶複合物技術和Vector Red染色基質(Vector Laboratories,伯靈格姆,CA)來顯示抗體分佈,然後用苯胺藍進行反染色。通過CD31的積極染色來識別毛細血管。隨機選擇每個組織製備的10個不同的視野,並計算可見的毛細血管。毛細血管密度表示為每平方毫米的毛細血管數量。為了評估缺血肌肉中的局部氧化應激水平,使用了抗硝酪氨酸的抗體(Upstate,Lake Placid,NY,USA)。
流式細胞計數
為了研究遠紅外(IFR)治療對組織缺血反應中EPC(內皮祖細胞)動員的影響,使用螢光活性細胞分類(FACS)Caliber流式細胞計數儀(Becton Dickinson,San Jose,CA,USA)來評估EPC的動員[18]。將100微升外周血與螢光異硫氰酸鹽(FITC)抗小鼠Sca-1(eBioscience,聖地亞哥,CA,USA)和藻紅素(PE)抗小鼠Flk-1(VEGFR-2,eBioscience)抗體孵育。同型同源抗體用作對照(Becton Dickinson,Franklin Lakes,NJ,USA)。孵育30分鐘後,細胞被溶解(PharmLyse; BD Pharmingen),用磷酸鹽緩沖鹽水(PBS)洗滌,並固定在2%的對甲醛中以供分析。每次分析包括100,000個事件。流通的EPC被認為來自單核細胞群,並且通過Sca-1和Flk-1的雙正門檻。
骨髓移植模型
8週齡的野生型受體小鼠被致命性輻射,總劑量為9.0 Gy[18, 20]。作為供體的是全面表達增強型GFP(eGFP)的eGFP轉基因小鼠(FVB背景)(Level Biotechnology Inc.,台北,台灣)。被輻射後,受體小鼠通過尾靜脈注射接受來自eGFP小鼠的未分級骨髓細胞(5×10^6個)。骨髓移植兩個月後,用STZ每日靜脈注射的方法誘導嵌合小鼠患糖尿病,如之前所述。所有小鼠都接受了後肢缺血手術(每組n = 6)。由流式細胞計數測量的eGFP陽性骨髓細胞再生為95%。在骨髓重構和STZ誘導的糖尿病小鼠進行後肢缺血手術兩週後,收集組織進行共軛焦免疫螢光和組織學分析。用針對eGFP(Chemicon)和CD31(BD PharMingen)的抗體染色骨髓源性EPC。通過計數eGFP+CD31+雙陽性細胞(黃色)在至少6個不同動物的不同橫截面下的高倍場(HPF,×100)下,估計EPC密度。
EPC在裸鼠中的移植
6–8週齡的無胸腺裸鼠由台灣國家實驗動物中心購得。然後,將裸鼠隨機分配到5個治療組(每組n = 6),分別進行肌肉內注射,包括正常生理鹽水、健康EPC、用高葡萄糖處理4天的EPC(EPC-HG)、用高葡萄糖和IFR處理30分鐘的EPC(EPC在高葡萄糖條件下處理4天後的24小時)(EPC-HG + IFR),以及用高葡萄糖處理並轉染eNOS siRNA(Santa Cruz-Biotechnology Inc.,CA,USA)和IFR處理30分鐘的EPC(EPC-HG + eNOS siRNA + IFR)。EPCs標記有螢光碳氰基染料1,1′-二十八烷基-1至3,3,3′,3′-四甲基印青碳氰基氯酸鹽(DiI)染料(Molecular Probes)[21]。單側後肢缺血手術後24小時進行肌肉內注射。總體積200微升的正常生理鹽水或5 × 10^6個EPC被注射到動脈閉塞部位以下的缺血後肢的6個部位。三個腹側注射位於靠近拈肌和半膜肌的上肢。其餘3個注射則注入涉及腓腸肌和屈指深肌的腹側下肢。為了達到最大的實驗一致性,移植的EPC均來自相同的捐贈者,並在平行實驗中使用。
人類EPC的分離和培養
從健康年輕成年志願者身上獲取外周血樣本(20毫升),並通過與Histopaque-1077(1.077 g/ml,Sigma,聖路易斯,MO,USA)的密度梯度離心分離總單個核細胞(MNCs)[20]。簡而言之,MNCs(5×10^6)被植入2毫升內皮生長培養基(EGM-2 MV Cambrex,East Rutherford,NJ,USA)中,補充物置於纖連蛋白塗佈的6孔板上。經過4天的培養後,更換培養基並去除非黏附細胞;附著的早期EPC呈紡錘形。允許一定數量的早期EPC生長為ECFCs(形成內皮細胞的細胞群),這些細胞群在MNC培養開始後的2-4週內出現。ECFCs在密度處呈現鵝卵石形態和成熟內皮細胞的單層生長模式[22]。在本研究中,收集ECFCs並用於所有測試。
ECFC表徵
ECFCs被表徵為黏附細胞,其呈陽性反應,對內皮細胞和造血幹細胞表面標記,如先前所述[20]。通過免疫螢光染色來表徵ECFCs,用於表達VE-cadherin,血小板/內皮細胞粘附分子-1(PECAM-1,CD-31),和CD34,KDR,AC133和eNOS(Santa Cruz)。螢光圖像在激光掃描共焦顯微鏡下記錄。
測量活性氧物種(ROS)產生和硝酸鹽水平的測量
利用2′,7′-二氯螢光素醋酸酯(DCFH-DA,Molecular Probes)作為探針測定IFR治療對ECFCs中ROS產生的影響,用以檢測H2O2的存在。21 ECFCs(每孔10^4細胞)在96孔板中孵育於高葡萄糖培養基中4天。IFR輻射治療後,細胞孵育於20μmol/L DCFH-DA中45分鐘。使用Griess試劑[1%磺胺和0.1% N-(1-萘基)乙二胺在2%磷酸中]測量調節培養基中的硝酸鹽水平。使用螢光微板讀取器在485nm激發和530nm發射下評估螢光強度(相對螢光單位)。
EPC衰老測試
使用衰老細胞染色劑組(Sigma)確定細胞老化。簡而言之,用PBS洗滌後,ECFCs在PBS中的2%甲醛和0.2%戊二醛中固定6分鐘,然後在新鮮X-半乳糖苷酶染色溶液(1 mg/ml X-半乳糖苷,5 mM六氰鉀鐵鹽,和2 mM MgCl2; pH6)中在37°C下無CO2孵育12小時。染色後,計數綠色染色細胞和全部細胞,並計算β-半乳糖苷酶陽性細胞的百分比[20]。
EPC遷移測試
通過修改的Boyden室測試(Transwell,Costar)[20]評估ECFCs的遷移功能。簡單來說,如上所述,分離的ECFCs與胰蛋白酶/EDTA分離,然後將4×10^4 ECFCs放置在帶有聚碳酸酯膜(8-μm孔)的24孔transwell板的上室中,該膜含有無血清內皮生長培養基。VEGF(50 ng/ml)被添加到放置在下室的培養基中。孵育24小時後,用PBS簡單洗滌膜並用4%順醛固定。上膜側輕輕擦拭棉球。使用若丹寧溶液染色膜並小心取出。通過計數6個隨機高倍(×100)顯微鏡視野中的遷移細胞來評估ECFCs的遷移程度。
Western blotting分析
ECFCs在緩衝液中(62.5 mM Tris–HCl,2% SDS,10%甘油,0.5 mM PMSF,2 μg/ml aprotinin,pepstatin和leupeptin)中裂解,蛋白質裂解物經過SDS-PAGE,然後電轉印到PVDF膜上[12]。膜用單克隆抗體探測磷酸化內皮NO合成酶(p-eNOS),eNOS,Akt,p-Akt,VEGF,p-ERK,p-38 MAPK,HO-1(Cell Signaling)和β-actin(Sigma)。帶子通過化學發光檢測試劑可見。密度計量分析使用ImageQuant(Promega)軟件。
統計分析
結果以平均值 ± 標準誤差(SEM)給出。統計分析采用無配對的學生t檢驗或方差分析,然后進行Scheffe多重比較事後檢驗。數據使用SPSS軟件(版本14; SPSS,芝加哥,IL)進行分析。 p值小於0.05被視為統計學上顯著。
結果
IFR治療促進糖尿病小鼠的血流恢復
局部IFR治療於STZ誘導的糖尿病小鼠手術後2週內每天兩次進行30分鐘,而野生型和糖尿病對照小鼠則放置在34°C的加熱板上每天兩次加熱30分鐘,以避免本研究中的熱效應。如圖1A所示,經雷射多普勒成像測定,與野生型小鼠相比,未經IFR治療的STZ誘導糖尿病小鼠在缺血手術後的血流恢復延遲。同時,重複的IFR治療在STZ誘導的糖尿病小鼠中顯著提高了血流恢復率48% (每組n = 6)。但是,用eNOS抑制劑NG-硝基-L-精氨酸甲酯 (L-NAME, 飲用水中1 mg/ml) 治療後,局部IFR輻射的好處明顯被消除。與雷射多普勒成像的測量結果一致,抗-CD31免疫染色揭示重複的FIR輻射增加了STZ誘導的糖尿病小鼠缺血肌肉中可檢測到的微血管數量 (對照組對IFR:38.8 ± 1.8對48.7 ± 2.4/HPF, p = 0.008) (圖1B)。但是,L-NAME的給予消除了IFR輻射的好處 (可檢測到的微血管,IFR對IFR + L-NAME:48.7 ± 2.4對34.8 ± 1.7/HPF, p = 0.001)。
圖1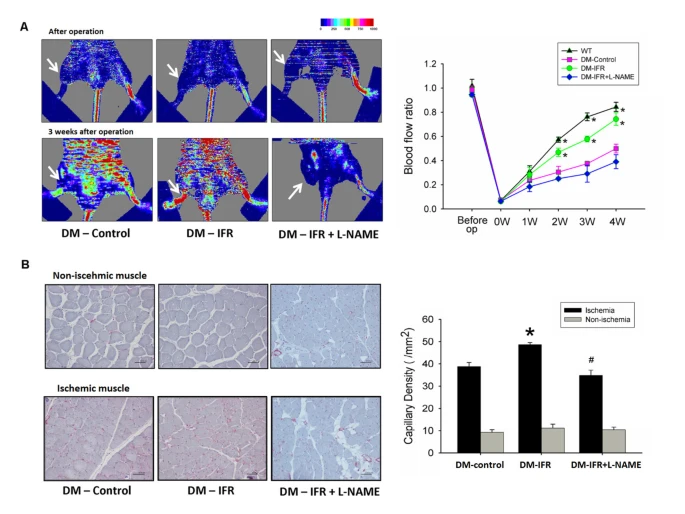{kind=link}
遠紅外線(IFR)治療對STZ誘導的糖尿病小鼠的血流恢復和新血管形成的影響。
(A) 野生型小鼠、對照組(載體)、IFR治療組和IFR + NG-硝基-L-精氨酸甲酯(L-NAME)小鼠手術前及後肢缺血手術4週後的雷射多普勒測量的代表結果。顏色尺度從最小(深藍色)到最大(紅色)值顯示血流變化。箭頭指示後肢缺血手術後的缺血(右)肢。不同組別中的多普勒灌注比率(缺血/非缺血後肢)隨著時間的變化。L-NAME在飲用水中的給予消除了IFR治療對糖尿病小鼠的有益效果。(*p < 0.05與DM-對照相比;#p < 0.05與DM-FIR相比;n = 6)
(B) 手術後3週犧牲小鼠,並使用抗-CD31免疫染色在缺血肌肉中顯現微血管。結果是平均值 ± 標準誤差(SEM)。(*p < 0.05與DM-對照相比;#p < 0.05與DM-IFR相比;n = 6)
IFR輻射對缺血肢體的氧化壓力、EPC動員和歸巢過程的影響
為了進一步評估IFR治療對缺血肌肉的氧化壓力的影響,進行了對硝基酪氨酸的免疫染色。如圖2A所示,通過硝基酪氨酸染色評估,在接受IFR治療的糖尿病小鼠的缺血肌肉中,氧化壓力水平顯著降低。
圖2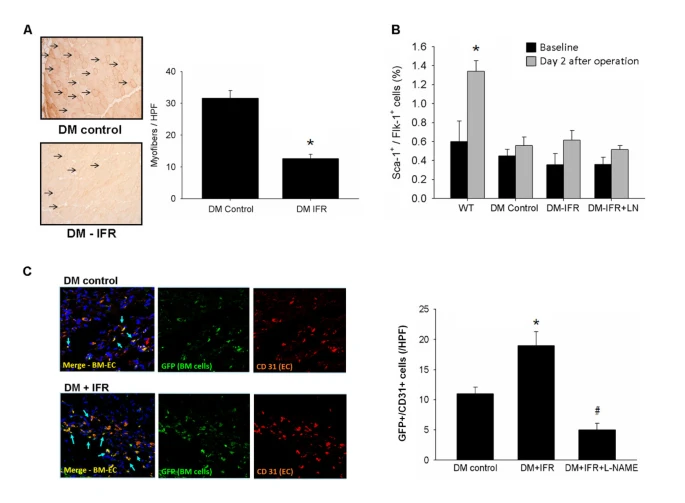{kind=link}
FR輻射對STZ誘導的糖尿病小鼠後肢缺血後的氧化壓力、EPC動員和組織定位的影響。
(A) IFR對STZ誘導的糖尿病小鼠缺血肌肉的氧化壓力的影響。第21天提取的缺血肌肉中的硝基酪氨酸(每組n = 4)免疫染色在對照組(載體)和接受IFR輻射的小鼠中。(*p < 0.05與DM-對照組相比)
(B) 組織缺血後的EPCs(定義為Sca-1+/Flk-1+細胞)動員在接受載體、IFR或IFR + L-NAME的STZ誘導的糖尿病小鼠中由流式細胞儀測定。(*p < 0.05與WT-基線相比;每組n = 6)
(C) 在接受eGFP小鼠骨髓細胞的FVB小鼠中產生了STZ誘導的糖尿病。通過免疫螢光染色,IFR組的STZ誘導的糖尿病小鼠在缺血肌肉中比載體組有更多的GFP+/CD31+雙陽性細胞。(*p < 0.05與DM-對照相比;#p < 0.05與DM-FIR相比;n = 6)
為了調查重複的IFR輻射對於組織缺血引起的EPC動員的影響,通過流式細胞儀在STZ誘導的糖尿病小鼠(每組n = 6)的外周血中確定了Sca-1+/Flk-1+細胞的水平。在野生型小鼠中,EPCs的動員在組織缺血後增強(基線與手術後2天相比:0.60 ± 0.22對1.34 ± 0.11%,p = 0.016)。然而,在STZ誘導的糖尿病小鼠中,在後肢缺血後,外周血中EPCs的動員受到損害(基線對手術後2天:0.45 ± 0.07對0.56 ± 0.09%,p = 0.374)。如圖1所示,局部IFR輻射的給予並未增加在後肢缺血手術後STZ誘導的糖尿病小鼠中的EPC動員(基線對手術後2天:0.36 ± 0.12對0.61 ± 0.10%,p = 0.140)。
為了測試重複的IFR輻射對骨髓源性EPC歸巢和分化為內皮細胞的影響,STZ誘導的糖尿病在接受eGFP小鼠骨髓細胞的FVB小鼠中產生。通過免疫螢光染色,IFR組的STZ誘導的糖尿病小鼠在缺血肌肉中有比對照組更多的GFP+/CD31+雙陽性細胞(對照與IFR相比:11.2 ± 1.1對18.8 ± 2.0/HPF,p = 0.007,n = 6;圖2)。此外,eNOS抑製劑的給予顯著減少了IFR輻射對EPC歸巢的影響。這些結果表明,重複的IFR治療在組織缺血後並未促進EPC的動員,但可能增加外周血中EPCs對缺血組織的歸巢。
人類ECFCs的特性
如之前描述的[22],ECFCs是從健康的年輕成人志願者的外周血MNCs中分離出來的。最初播種在纖維連接蛋白塗層孔上的外周血MNCs呈圓形(圖3A)。當第4天更換培養基後,附著的MNCs呈現為拉長的紡錘形狀(圖3B)。ECFCs呈鵝卵石狀形態,類似成熟的內皮細胞,成為了密集生長(圖3C)。ECFCs的特性是通過免疫組織化學染色進行的,大多數細胞表達成熟的內皮標記,包括VE-cadherin、PECAM-1 (CD31)、CD34、KDR、AC133 和 eNOS(圖3),這些被認為是晚期EPCs的關鍵標記。
圖3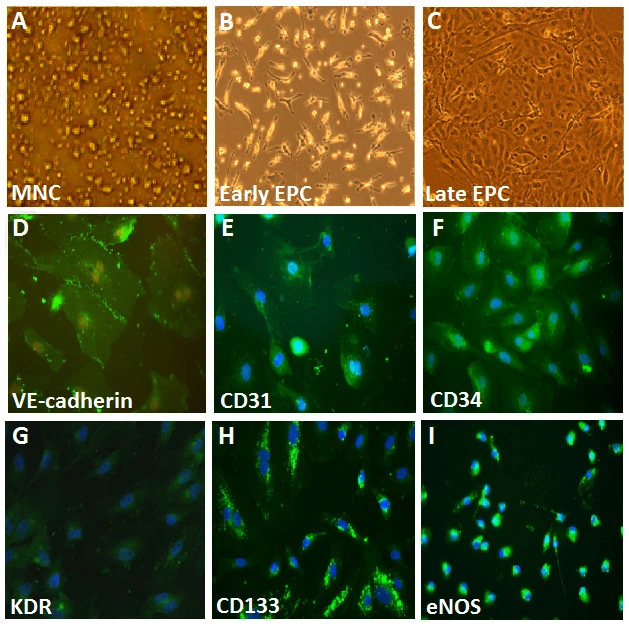{kind=link}
人類外周血內皮前驅細胞(EPCs)的形態和特性。
(A) 第一天,外周血單核細胞(MNCs)被播種在纖維連接蛋白塗層的培養盤上。
(B) 播種後的四天,附著的初級EPCs呈紡錘形狀。
(C) 播種後的三週,選擇的ECFCs呈鵝卵石狀形態,被重新播種,並生長到滿佈。
(D-I) ECFC的特性是通過免疫組織化學染色來進行的。大部分的EPC表達內皮和造血幹細胞標記,如VE-cadherin、PECAM-1 (CD31)、CD34、KDR、AC133和eNOS,這些被認為是EPCs的關鍵標記。細胞核(藍色)用4′,6-二胺基-2-苯基吲哚(DAPI)進行對照染色。
IFR 降低反應性氧化應激並在高糖環境中增強EPC增殖,NO產生
高糖顯著增加了由相對DCFH-DA螢光強度確定的H2O2產生,且IFR輻射(10–60 分鐘)顯著抑制了在ECFCs培養中由高糖誘導的ROS指數(圖4A)。
圖4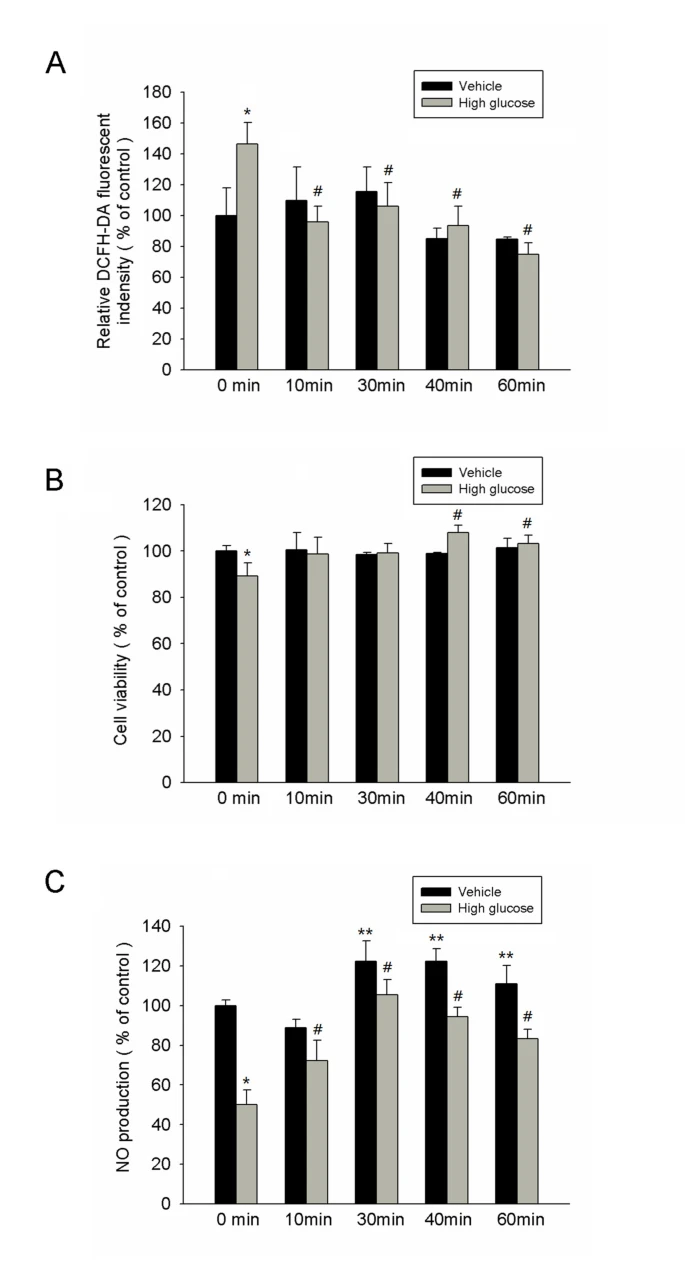{kind=link}
IFR 療法降低了反應性氧化應激,恢復了EPC增殖,並在高糖條件下增加了NO產量。
(A) 高糖顯著增加了由相對DCFH-DA螢光強度確定的H2O2產生,而使用IFR療法抑制了在EPCs培養中由高糖誘導的反應性氧化應激(ROS)指數。 (*p < 0.05 與控制-0分鐘比較; #p < 0.05 與高糖-0分鐘比較)
(B) 通過MTT試驗分析了IFR輻射對EPCs增殖的影響。 (*p < 0.05 與控制-0分鐘比較; #p < 0.05 與高糖-0分鐘比較)
(C) 通過Griess試劑測量了培養基中的硝酸鹽產量(作為NO含量)。高糖抑制了培養的晚期EPCs中的NO產量。在培養4天後,IFR輻射在有或沒有高糖條件下增加了NO產量。(*p < 0.05 與控制-0分鐘比較; #p < 0.05 與高糖-0分鐘比較; **p < 0.05 與控制-0分鐘比較; 每個實驗 n = 4)。
如圖4B所示,用高糖培養基孵育4天顯著減少了EPC的增殖(控制組與高糖組比較,100 ± 2與88 ± 5, p = 0.026)。然而,對EPCs施加IFR輻射40分鐘後,再在高糖條件下處理,可以顯著逆轉高糖對EPC增殖的減少(高糖組與高糖+ IFR 40分鐘組比較,88 ± 5對109 ± 2, p = 0.006)。
高糖會損害eNOS的活化並減少培養的EPCs中的NO生物可利用性 [7]。因此,我們測試了IFR輻射對高糖處理的EPCs的效果,以確定IFR是否能夠恢復EPCs中受損的NO產量。在25 mM高糖培養基中孵育4天後,培養基中的NO產量顯著下降(圖4C)。但是,對EPCs進行30分鐘的IFR輻射處理後,再在高糖條件下的處理可以顯著上調高糖對No產量的損害。
我們研究了IFR輻射對高糖處理的EPCs的影響,以確定IFR治療是否可以恢復EPCs中受損的eNOS啟動。
對培養的ECFCs進行10分鐘、30分鐘和40分鐘的IFR輻射後,再用高糖條件處理EPCs,顯著上調了高糖損害的eNOS產量和eNOS活性(p-eNOS/總eNOS)。使用IFR治療30分鐘、40分鐘和60分鐘也上調了Akt的啟動(p-Akt/總Akt)。此外,IFR輻射還促進了高糖條件下的VEGF產量、p-ERK和p-38 MAPK,但HO-1則沒有(圖5)。這些發現表明,使用IFR輻射可能會增強EPCs對高糖刺激的反應,提高eNOS、Akt、p-ERK、p-38 MAPK和VEGF的活性。
圖5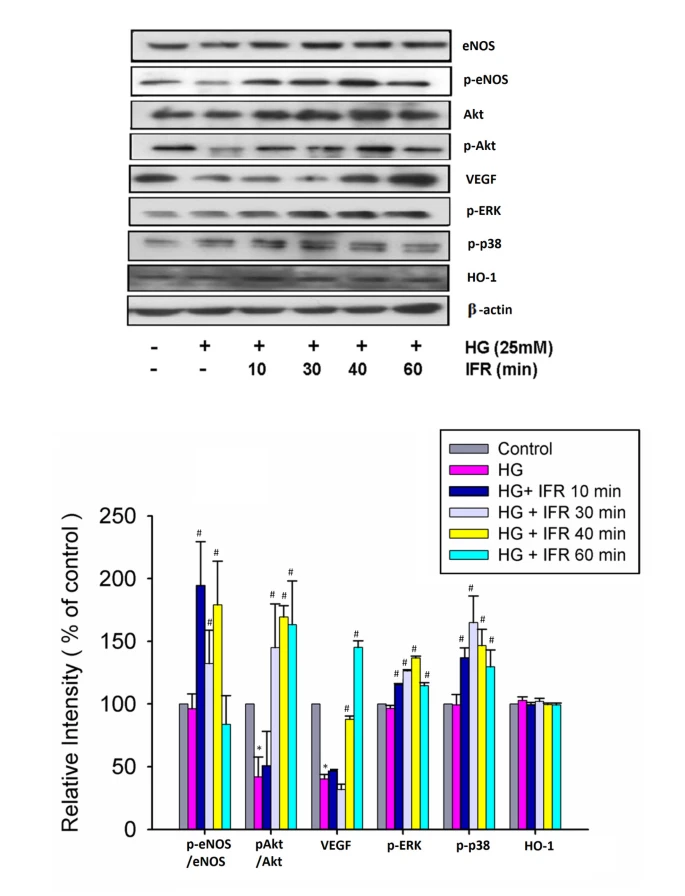{kind=link}
IFR輻射對培養的EPCs中的eNOS、Akt、p-ERK、p-38 MAPK、VEGF和HO-1產量的影響。對培養的ECFCs進行10分鐘、30分鐘和40分鐘的IFR輻射後,再用高糖條件處理EPCs,顯著上調了高糖損害的eNOS產量和eNOS活性(p-eNOS/總eNOS)。進行IFR治療30分鐘、40分鐘和60分鐘也上調了Akt的啟動(p-Akt/總Akt)。此外,IFR輻射還促進了高糖條件下的VEGF產量、p-ERK和p-38 MAPK,但HO-1則沒有。每個條形圖顯示了四個獨立實驗的彙總數據,使用光譜密度法進行正常化後。數據是平均值±SEM;每個實驗中n=4。(*p < 0.05與對照組相比;#p < 0.05與HG組相比)。
IFR輻射對培養的EPCs中的eNOS、Akt、p-ERK、p-38 MAPK、VEGF和HO-1產量的影響。
對培養的ECFCs進行10分鐘、30分鐘和40分鐘的IFR輻射後,再用高糖條件處理EPCs,顯著上調了高糖損害的eNOS產量和eNOS活性(p-eNOS/總eNOS)。進行IFR治療30分鐘、40分鐘和60分鐘也上調了Akt的啟動(p-Akt/總Akt)。此外,IFR輻射還促進了高糖條件下的VEGF產量、p-ERK和p-38 MAPK,但HO-1則沒有。每個條形圖顯示了四個獨立實驗的彙總數據,使用光譜密度法進行正常化後。數據是平均值±SEM;每個實驗中n=4。(*p < 0.05與對照組相比;#p < 0.05與HG組相比)。
圖6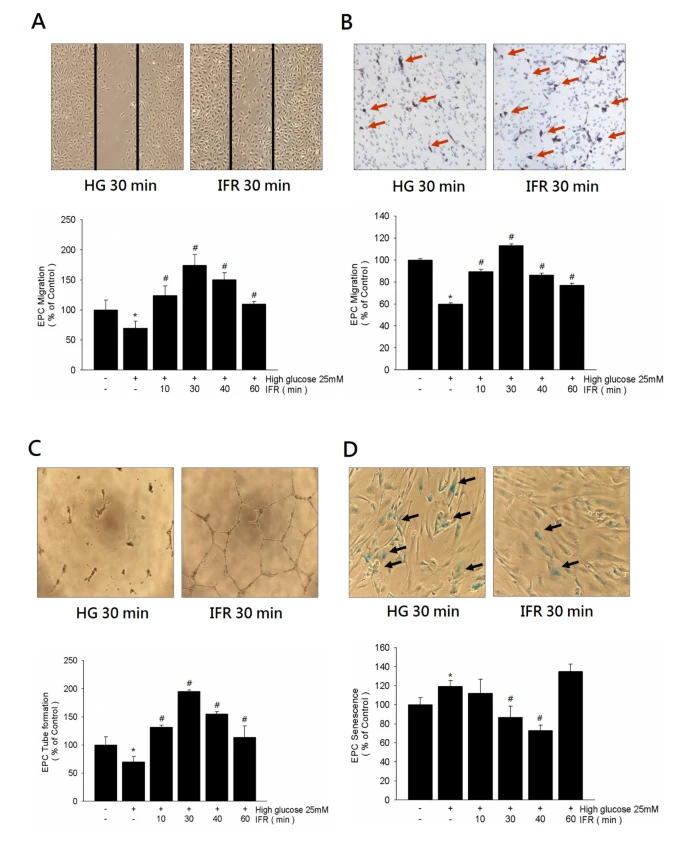{kind=link}
IFR療法對體外EPC遷移、管腔形成和老化的影響。
(A,B) 刮痕試驗和改良的Boyden室分析用於評估高糖條件下ECFCs的遷移功能。使用VEGF作為趨化因子的Boyden室分析用於評估IFR輻射對EPC遷移的影響。
(C) 針對晚期ECFCs的體外血管新生分析使用了ECMatrix凝膠。顯示了體外血管新生的代表性照片。細胞用結晶紫染色,並使用計算機軟件比較由細胞形成的完整管腔的總面積的平均值。
(D) 為了確定細胞老化的開始,酸性ß-半乳糖苷酶被用作酸化的生化標記,這是典型的ECFCs老化。數據是平均值±SEM;每個實驗n=4。
培養4天後,EPCs在ECMatrix凝膠上的管腔形成能力在高糖存在下明顯減少,與對照組相比。而對EPCs進行30分鐘的IFR輻射治療,接著在高糖條件下進行EPCs治療,則改善了此高糖抑制的EPCs管腔形成(100 ± 15 vs. 195 ± 3 cells/HPF, p = 0.003;圖6C)。
與對照組相比,使用高糖孵化的EPCs顯著增加了與衰老相關的ß-半乳糖苷酶陽性EPCs的百分比(100 ± 6對119 ± 5%,p = 0.032)。30分鐘的IFR輻射治療後,接著在高糖培養基中對EPCs進行治療,明顯減少了與衰老相關的ß-半乳糖苷酶陽性EPCs的百分比(高糖對高糖+IFR, 119 ± 5對87 ± 9%,p = 0.013;圖6D)。
經EPC處理的IFR輻射移植改善了後肢血流灌流情況。
接受正常生理鹽水的動物,在整個研究期間血流保持穩定,約為非缺血肢體的42%(手術後四周的血流量為42 ± 8%)。相比之下,接受EPC和EPC在高葡萄糖培養基中接受IFR照射(30分鐘)移植的小鼠(EPC-HG + IFR),但不接受IFR療法的高葡萄糖培養基中的EPC(EPC-HG)以及接受高葡萄糖和IFR和eNOS siRNA處理的EPC(EPC-HG + eNOS siRNA + IFR)在EPC植入後三周已經顯著改善了血流量(兩者均p < 0.05;圖7)。與激光多普勒影像測量一致,抗CD31免疫染色顯示,接受EPC和EPC在高葡萄糖條件下接受IFR處理的移植小鼠,在缺血肌肉中可檢測到的毛細血管數量顯著增加,而與接受正常生理鹽水的小鼠相比(兩者均p < 0.05比較正常生理鹽水)。然而,eNOS siRNA處理消除了高葡萄糖培養基中IFR輻射的效益(IFR與IFR + eNOS siRNA:38.7 ± 4.7與21.3 ± 3.7 / HPF,p = 0.015,每組n = 6)。
圖7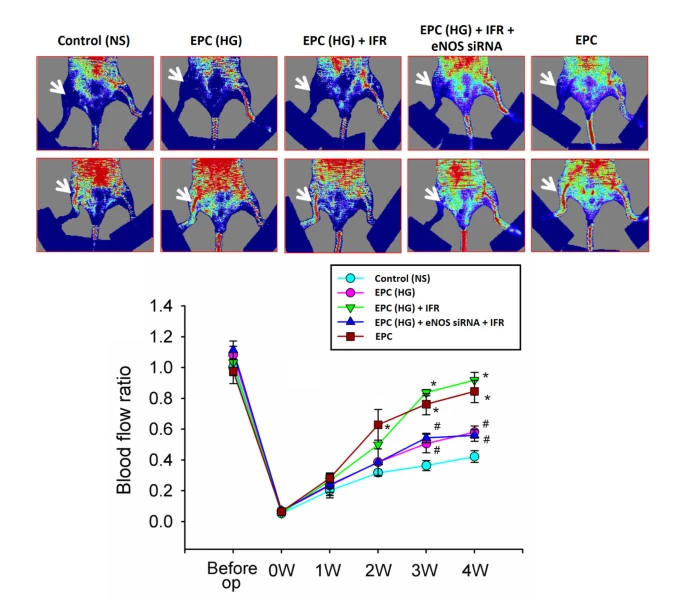{kind=link}
經EPC處理的IFR輻射移植改善了缺血後肢的血液灌流情況。 (A) 血液灌流量的代表性影像,由激光多普勒測量,以及在缺血後肢手術後立即及注射正常鹽水、EPC處理高葡萄糖(EPC-HG)、EPC處理高葡萄糖並進行IFR療法(EPC-HG + IFR)或EPC處理高葡萄糖和IFR和eNOS siRNA(EPC-HG + eNOS siRNA + IFR)後3週的血流量定量分析,以灌流率表示(缺血/對側非手術的後肢)。 (*p <0.05與對照組相比; #p <0.05與EPC-HG + IFR相比; n = 6)。
討論
本研究首次顯示IFR輻射對糖尿病小鼠的缺血誘導新血管形成具有有利效果。重複的IFR輻射增強了缺血後肢的血流恢復和新血管的形成,而這一有益效果可能源於通過減少缺血組織中的氧化應激來增強EPC歸巢過程,而不是促進EPC動員。此外,直接的IFR輻射可以減輕高糖誘導的氧化應激,減弱細胞衰老並改善EPC功能。接受用IFR輻射處理的EPC的小鼠在組織缺血後的血流恢復中表現出顯著的改善,與接受生理鹽水的小鼠相比。考慮到上述證據,我們的研究結果表明IFR輻射對組織缺血後血流恢復的直接有益效果,而且IFR輻射可以抵消糖尿病環境對改善EPC功能的不利影響,這可能為其對血管保護的潛在臨床影響提供了一些新的理由。
為響應組織缺血而改善的新血管形成是減少器官損傷的重要治療策略。有力的證據表明,成人的新血管形成不僅僅是內皮細胞(血管生成)增殖的結果,而且還涉及到血管生成過程中的循環EPCs。這些循環EPCs來自骨髓,並由組織缺血或外源性的細胞因子刺激,如VEGF和基質細胞衍生因子-1 (SDF-1) [23],內源性地被動員。
然而,患有糖尿病或心血管風險因素的患者被發現循環EPCs的數量和功能減少[8, 24, 25]。最近的研究指出,先進的糖化終產物(AGEs)促進EPCs的凋亡[26],長期暴露於高糖可能透過NO相關機制增強細胞衰老,並減少EPCs的細胞數量和功能能力[10]。EPCs的動員和分化由NO調節,骨髓表達的eNOS對幹細胞和前驅細胞的動員至關重要[27]。此外,內源性NOS抑制劑,如非對稱的二甲基精胺(ADMA),被證明抑制EPCs的分化和功能,並導致內皮功能受損[28]。這些發現為高血糖抑制EPC功能,以及與糖尿病相關的血管併發症提供了潛在的治療目標理由。
糖尿病患者經常遭受微血管或巨血管異常,包括視網膜病變、腎病、神經病變和加速的動脈粥樣硬化。從eNOS產生的NO生物可用性減少在動脈粥樣硬化的發展和進展中起到關鍵作用是明確的。在各種病理狀態下,如2型糖尿病,eNOS可能會失去功能或其表達可能會減少。內皮功能障礙與兒童肥胖相關,並與EPCs的數量和功能密切相關,綜合放學後的運動計劃被證明可以通過增強NO生物可用性來增加循環EPCs水平[29]。此外,由於自由基可以直接使NO失活,組織缺血可能會降低NO的生物可用性,從而增強氧化應激[30]。糖尿病患者的缺血肢體或心肌對缺血的血管生成反應不足可能導致膠原形成不良和嚴重的器官損傷。
IFR輻射是一種看不見的電磁波,其特徵波長在5.6至1000 μm之間,可以通過皮膚中的熱感受器感知為熱[11, 12]。IFR的技術已被廣泛應用於各種領域。IFR的熱效應導致血管擴張和增加組織血流。局部IFR治療可能允許多重能量傳輸深入皮下組織2到3公分,而不會像未過濾的熱輻射那樣刺激或過熱皮膚[31]。當陶瓷板和皮膚之間的距離>20公分時,皮膚溫度在使用FIR治療30到60分鐘期間穩定上升到大約38到39°C的高度[16]。因此,紅外線治療可以免於熱療的不利影響或副作用。在本研究中,野生型和糖尿病對照小鼠被放置在34°C的加熱板上,每天兩次,各30分鐘,以避免組間的熱效應。這些小鼠的直腸溫度也被評估,並顯示接受IFR治療的小鼠與未接受IFR輻射的小鼠相比沒有體溫上升。
除了熱效應外,越來越多的證據表明IFR治療的非熱效應通過NO相關途徑對心血管系統產生有益效果[31, 32]。在動物研究中,Akasaki及其同事證明了重複的IFR治療可以上調eNOS的表達並增強在單側後肢缺血的脂蛋白E缺乏小鼠模型中的血管生成[16]。在人體研究中,Imamura等人顯示,重複兩週的桑拿治療顯著改善了血管內皮功能,從而使得冠狀動脈風險因子患者中由4%增加到5.8%的流動介導的、內皮依賴性的上臂動脈擴張[14]。這些發現暗示IFR的非熱效應可能來自於上調eNOS活性和增強NO生物可用性。在這項研究中,我們證明重複的IFR治療可以激活eNOS和Akt,並上調ECFCs的遷移和管狀結構形成能力。Akt是PI 3-kinase的下游,能夠直接在Ser1179位置磷酸化eNOS,從而激活它。最近的證據也顯示,p-38 MAPK在糖尿病患者中由於高血糖而下調EPCs中起到了關鍵作用[33]。暴露於IFR的EPCs上調了p-38 MAPK和p-ERK的表達。這些發現與最近的研究相一致,該研究表明IFR輻射通過成熟內皮細胞上的MAP激酶依賴性途徑顯著促進了血管生成。在高糖環境下接受用IFR輻射處理的EPC的小鼠與接受普通鹽水的小鼠相比,在組織缺血後的血流恢復上顯示出顯著的改善。這些有益效果可能為糖尿病患者在關鍵性四肢缺血或臨床治療功能失常的EPCs之前提供了IFR治療的血管保護特性的一些新的理由。
結論
此研究首次提供證據,顯示IFR治療在STZ誘導的糖尿病小鼠中促進了旁路血流恢復和新的血管形成。這些有益效果可能源於增強EPC功能和歸巢過程,為糖尿病患者在關鍵性四肢缺血或進一步的細胞治療中IFR治療的血管保護特性提供了一些新的理由。
縮寫
EPCs:內皮前體細胞
NO:一氧化氮
DM:糖尿病
IFR:遠紅外線
STZ:鏈黴素
FACS:螢光活性細胞分選
FITC:異硫氰酸螢光素酯
PE:藍綠藻紅蛋白
PBS:磷酸緩衝鹽水
MNCs:單核細胞
ECFCs:形成內皮集落的細胞
PECAM-1:血小板/內皮細胞黏附分子-1
ROS:活性氧物種
DCFH-DA:2′,7′-二氯螢光素二醋酸酯
p-eNOS:磷酸化內皮NO合酶
L-NAME:NG-硝基-L-精胺甲酯
SDF-1:間質細胞衍生因子-1
AGEs:先進糖化終產物
ADMA:非對稱二甲基精胺。
縮寫
-
Folkman J: Seminars in Medicine of the Beth Israel Hospital, Boston. Clinical applications of research on angiogenesis. N Engl J Med. 1995, 333 (26): 1757-1763. 10.1056/NEJM199512283332608.
-
Freedman S, Isner JM: Therapeutic angiogenesis for ischemic cardiovascular disease. J Mol Cell Cardiol. 2001, 33 (3): 379-393. 10.1006/jmcc.2000.1329.
-
Rosengart TK, Lee LY, Patel SR, Sanborn TA, Parikh M, Bergman GW, Hachamovitch R, Szulc M, Kligfield PD, Okin PM, Hahn RT, Devereux RB, Post MR, Hackett NR, Foster T, Grasso TM, Lesser ML, Isom OW, Crystal RG: Angiogenesis gene therapy: phase I assessment of direct intramyocardial administration of an adenovirus vector expressing VEGF121 cDNA to individuals with clinically significant severe coronary artery disease. Circulation. 1999, 100 (5): 468-474. 10.1161/01.CIR.100.5.468.
-
Nathan DM, Cleary PA, Backlund JY, Genuth SM, Lachin JM, Orchard TJ, Raskin P, Zinman B: Diabetes Control and Complications Trial/Epidemiology of Diabetes Interventions and Complications (DCCT/EDIC) Study Research Group: Intensive diabetes treatment and cardiovascular disease in patients with type 1 diabetes. N Engl J Med. 2005, 353 (25): 2643-2653.
-
Adeghate E: Molecular and cellular basis of the etiology and management of diabetic cardiomyopathy: a short review. Mol Cell Biochem. 2004, 261 (1–2): 187-191.
-
Federman DG, Bravata DM, Kirsner RS: Peripheral arterial disease: systemic disease extending beyond the affected extremity. Geriatrics. 2004, 59 (4): 26.
-
Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM: Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997, 275 (5302): 964-967. 10.1126/science.275.5302.964.
-
Vasa M, Fichtlscherer S, Aicher A, Adler K, Urbich C, Martin H, Zeiher AM, Dimmeler S: Number and migratory activity of circulating endothelial progenitor cells inversely correlate with risk factors for coronary artery disease. Circ Res. 2001, 89 (1): E1-E7. 10.1161/hh1301.093953.
-
Chen YH, Lin SJ, Lin FY, Wu TC, Tsao CR, Huang PH, Liu PL, Chen YL, Chen JW: High glucose impairs early and late endothelial progenitor cells by modifying nitric oxide-related but not oxidative stress-mediated mechanisms. Diabetes. 2007, 56 (6): 1559-1568. 10.2337/db06-1103.
-
Yue WS, Lau KK, Siu CW, Wang M, Yan GH, Yiu KH, Tse HF: Impact of glycemic control on circulating endothelial progenitor cells and arterial stiffness in patients with type 2 diabetes mellitus. Cardiovasc Diabetol. 2011, 10: 113-10.1186/1475-2840-10-113.
-
Toyokawa H, Matsui Y, Uhara J: Promotive effects of far-infrared ray on full-thickness skin wound healing in rats. Exp Biol Med. 2003, 228 (6): 724-729.
-
Capon A, Mordon S: Can thermal lasers promote skin wound healing?. Am J Clin Dermatol. 2003, 4 (1): 1-12. 10.2165/00128071-200304010-00001.
-
Kihara T, Biro S, Ikeda Y: Effects of repeated sauna treatment on ventricular arrhythmias in patients with chronic heart failure. Circ J. 2004, 68 (12): 1146-1151. 10.1253/circj.68.1146.
-
Imamura M, Biro S, Kihara T: Repeated thermal therapy improves impaired vascular endothelial function in patients with coronary risk factors. J Am Coll Cardiol. 2001, 38 (4): 1083-1088. 10.1016/S0735-1097(01)01467-X.
-
Lin CC, Liu XM, Peyton K, Wang H, Yang WC, Lin SJ, Durante W: Far infrared therapy inhibits vascular endothelial inflammation via the induction of heme oxygenase-1. Arterioscler Thromb Vasc Biol. 2008, 28 (4): 739-745. 10.1161/ATVBAHA.107.160085.
-
Akasaki Y, Miyata M, Eto H, Shirasawa T, Hamada N, Ikeda Y, Biro S, Otsuji Y, Tei C: Repeated thermal therapy up-regulates endothelial nitric oxide synthase and augments angiogenesis in a mouse model of hindlimb ischemia. Circ J. 2006, 70 (4): 463-470. 10.1253/circj.70.463.
-
Yu SY, Chiu JH, Yang SD, Hsu YC, Lui WY, Wu CW: Biological effect of far-infrared therapy on increasing skin microcirculation in rats. Photodermatol Photoimmunol Photomed. 2006, 22 (2): 78-86. 10.1111/j.1600-0781.2006.00208.x.
-
Huang PH, Lin CP, Wang CH, Chiang CH, Tsai HY, Chen JS, Lin FY, Leu HB, Wu TC, Chen JW, Lin SJ: Niacin improves ischemia-induced neovascularization in diabetic mice by enhancement of endothelial progenitor cell functions independent of changes in plasma lipids. Angiogenesis. 2012, 15 (3): 377-389. 10.1007/s10456-012-9267-z.
-
Kilkenny C, Browne WJ, Cuthill IC, Emerson M, Altman DG: Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8 (6): 1000412-10.1371/journal.pbio.1000412.
-
Huang PH, Chen YH, Wang CH, Chen JS, Tsai HY, Lin FY, Lo WY, Wu TC, Sata M, Chen JW, Lin SJ: Matrix metalloproteinase-9 is essential for ischemia-induced neovascularization by modulating bone marrow-derived endothelial progenitor cells. Arterioscler Thromb Vasc Biol. 2009, 29 (8): 1179-1184. 10.1161/ATVBAHA.109.189175.
-
Huang PH, Chen JS, Tsai HY, Chen YH, Lin FY, Leu HB, Wu TC, Lin SJ, Chen JW: Globular adiponectin improves high glucose-suppressed endothelial progenitor cell function through endothelial nitric oxide synthase dependent mechanisms. J Mol Cell Cardiol. 2011, 51 (1): 109-119. 10.1016/j.yjmcc.2011.03.008.
-
Huang PH, Huang SS, Chen YH, Lin CP, Chiang KH, Chen JS, Tsai HY, Lin FY, Chen JW, Lin SJ: Increased circulating CD31+/annexin V + apoptotic microparticles and decreased circulating endothelial progenitor cell levels in hypertensive patients with microalbuminuria. J Hypertens. 2010, 28 (8): 1655-1665. 10.1097/HJH.0b013e32833a4d0a.
-
Takahashi T, Kalka C, Masuda H, Chen D, Silver M, Kearney M, Magner M, Isner JM, Asahara T: Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med. 1999, 5 (4): 434-438. 10.1038/7434.
-
Fadini GP, de Kreutzenberg S, Albiero M, Coracina A, Pagnin E, Baesso I, Cignarella A, Bolego C, Plebani M, Nardelli GB, Sartore S, Agostini C, Avogaro A: Gender differences in endothelial progenitor cells and cardiovascular risk profile: the role of female estrogens. Arterioscler Thromb Vasc Biol. 2008, 28 (5): 997-1004. 10.1161/ATVBAHA.107.159558.
-
Palombo C, Kozakova M, Morizzo C, Gnesi L, Barsotti MC, Spontoni P, Massart F, Salvi P, Balbarini A, Saggese G, Di Stefano R, Federico G: Circulating endothelial progenitor cells and large artery structure and function in young subjects with uncomplicated type 1 diabetes. Cardiovasc Diabetol. 2011, 10: 88-10.1186/1475-2840-10-88.
-
Li H, Zhang X, Guan X, Cui X, Wang Y, Chu H, Cheng M: Advanced glycation end products impair the migration, adhesion and secretion potentials of late endothelial progenitor cells. Cardiovasc Diabetol. 2012, 11: 46-10.1186/1475-2840-11-46.
-
Aicher A, Heeschen C, Mildner-Rihm C, Urbich C, Ihling C, Technau-Ihling K, Zeiher AM, Dimmeler S: Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat Med. 2003, 9 (11): 1370-1376. 10.1038/nm948.
-
Thum T, Tsikas D, Stein S, Schultheiss M, Eigenthaler M, Anker SD, Poole-Wilson PA, Ertl G, Bauersachs J: Suppression of endothelial progenitor cells in human coronary artery disease by the endogenous nitric oxide synthase inhibitor asymmetric dimethylarginine. J Am Coll Cardiol. 2005, 46 (9): 1693-1701. 10.1016/j.jacc.2005.04.066.
-
Park JH, Miyashita M, Kwon YC, Park HT, Kim EH, Park JK, Park KB, Yoon SR, Chung JW, Nakamura Y, Park SK: A 12-week after-school physical activity programme improves endothelial cell function in overweight and obese children: a randomised controlled study. BMC Pediatr. 2012, 12 (1): 111-10.1186/1471-2431-12-111.
-
Gryglewski RJ, Palmer RM, Moncada S: Superoxide anion is involved in the breakdown of endothelium-derived vascular relaxing factor. Nature. 1986, 320 (6061): 454-456. 10.1038/320454a0.
-
Hartel M, Hoffmann G, Wente MN, Martignoni ME, Buchler MW, Friess H: Randomized clinical trial of the influence of local water-filtered infrared A irradiation on wound healing after abdominal surgery. Br J Surg. 2006, 93 (8): 952-960. 10.1002/bjs.5429.
-
Hsu YH, Chen YC, Chen TH, Sue YM, Cheng TH, Chen JR, Chen CH: Far-infrared therapy induces the nuclear translocation of PLZF which inhibits VEGF-induced proliferation in human umbilical vein endothelial cells. PLoS One. 2012, 7 (1): e30674-10.1371/journal.pone.0030674.
-
Seeger FH, Haendeler J, Walter DH, Rochwalsky U, Reinhold J, Urbich C, Rössig L, Corbaz A, Chvatchko Y, Zeiher AM, Dimmeler S: p38 mitogen-activated protein kinase downregulates endothelial progenitor cells. Circulation. 2005, 111 (9): 1184-1191. 10.1161/01.CIR.0000157156.85397.A1.